04-05-2022
Door Andrew M. Allen
Populatiemodellen helpen bij het vaststellen of een populatie groeit of afneemt, d.w.z. de populatiedynamiek - en in termen van natuurbescherming, hoe verbeteringen in reproductie of overleving de populatiegroei kunnen beïnvloeden. Voor een langlevende soort als de scholekster is het ontwikkelen van populatiemodellen die de populatietrends nauwkeurig beschrijven geen eenvoudige opgave. In tegenstelling tot kleinere vogels beginnen scholeksters aanzienlijk later met de voortplanting (d.w.z. ze beginnen zich pas op of na hun derde kalenderjaar te reproduceren). Verder hebben scholeksters een hoge levensverwachting en vertonen daarom waarschijnlijk leeftijdsafhankelijke overlevings- en reproductiekansen, en er is grote individuele variatie in bijvoorbeeld waar ze broeden (kust, grasland , landbouw, stedelijke gebieden), voedselspecialisaties (bijv. wormen of schelpdieren) en de data waarop individuen hun nest beginnen (bijv. vroeg versus laat in het broedseisoen). Gelukkig is er grote vooruitgang geboekt in de statistische berekeningen voor populatiemodellering. Een belangrijke vooruitgang is de ontwikkeling van een methode genaamd integrale projectiemodellen (IPM's), waarmee wetenschappers individuele variatie in hun modellen kunnen opnemen, en niet alleen in categorieën zoals geslacht of leeftijdsklasse , maar zelfs variatie die continu kan worden gemeten, bijvoorbeeld de dag van het jaar dat scholeksters beginnen te broeden. Deze modellen geven nieuwe inzichten in de populatiedynamiek van soorten, en we gebruiken deze methode om de scholeksterpopulatie op Schiermonnikoog te bestuderen. Een belangrijke vraag is hoe we populatieherstel op Schiermonnikoog en uiteindelijk in heel Nederland het beste kunnen realiseren.
De scholeksterpopulatie op Schiermonnikoog wordt al bijna vier decennia gevolgd, maar neemt al bijna drie decennia af. Al die tijd is de populatie in detail bestudeerd, met goede schattingen van reproductie, inclusief nestsucces en overleving van de kuikens. De meeste vogels zijn voorzien van een kleurring, wat betekent dat we individuen kunnen volgen en demografische parameters zoals overleving kunnen inschatten en hoe broedstatus of legdatum tussen jaren kunnen verschillen.
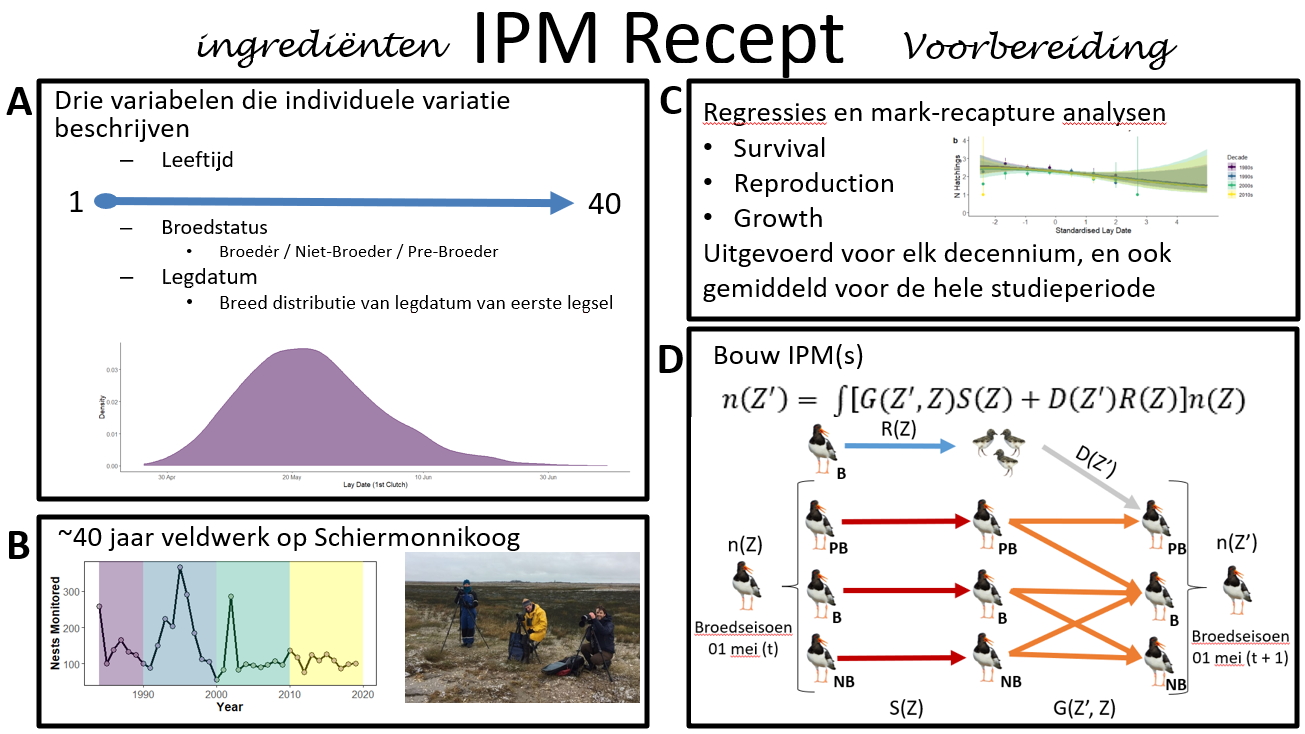
Figuur 1. Recept voor het maken van een integraal projectiemodel met de ingrediënten (a,b) voor het meten van individuele variatie in overleving, reproductie en eigenschappen, en het volgen van individuen over een lange tijdsperiode door intensief veldwerk, en de voorbereiding (c,d) van het analyseren van alle relaties voorafgaand aan het ontwerpen en maken van het populatiemodel volgens de levensgeschiedeniskenmerken van de onderzoekspopulatie.
Wat zijn de ingrediënten voor het integrale projectiemodel? E erst moeten we de individuen in de populatie nauwkeurig beschrijven en we doen dit met behulp van drie variabelen (Figuur 1a), namelijk i) Leeftijd - die bestaat uit 40 leeftijdscategorieën van 1 tot 40 (ongeveer de maximale levensduur van een scholekster), ii) Broedstatus, omdat alleen broedvogels zich kunnen voortplanten, overleven varieert tussen broedvogels en niet-broedvogels, en de kans om volgend jaar te broeden afhankelijk is van de huidige broedstatus, en iii) legdatum, omdat we grote verschillen tussen individuen in legdatum hebben waargenomen (zie inzetafbeelding voor verdeling van legdatums in figuur 1a) en reproductief succes is sterk afhangt van de legdatum. Om vast te kunnen stellen hoe overleving, voortplanting en broedkans samenhangen met leeftijd, broedstatus en legdatum, is een grote inspanning nodig om de populatie te monitoren voor een soort als de scholekster die meer dan 40 jaar kan leven. Dankzij het harde werk van onderzoekers en vrijwilligers zijn er sinds 1983 jaarlijks meer dan 100 nesten op Schiermonnikoog gemonitord (Figuur 1b).
Om de IPM voor te bereiden, moet men vervolgens een aantal analyses uitvoeren (regressies en mark-recapture-analyses; figuur 1c) om de relatie tussen legdatum en reproductie, legdatum, broedstatus en leeftijd met overleving te identificeren, en hoe de broedkans varieert met huidige broedstatus en leeftijd. Enkele voorbeelden van de relaties zijn weergegeven in figuur 2, bijvoorbeeld de relatie tussen het aantal geproduceerde jongen (die het eerste jaar overleefde) en de legdatum, waarbij vogels met latere legdatums het duidelijk slechter doen dan vogels met vroegere legdatums, maar deze relatie is ook veranderd tussen 1980 en vandaag (Figuur 2a). Broeders hebben een hogere overlevingskans dan niet-broeders, maar er is geen relatie tussen overlevingskans van volwassen vogels en legdatum (Figuur 2b). Voor jonge vogels, hebben vlieggevlugde kuikens de laagste overleving (Age0), gevolgd door 2kj-vogels (Age1) en 3kj-vogels (Age2), en kuikens uit nesten met vroegere legdatums hebben een hogere overleving, vooral in het eerste jaar (Figuur 2c). Ten slotte zijn de broedkansen aanvankelijk laag voor jonge vogels (3j+) maar nemen ze toe met de leeftijd voordat ze weer afnemen naarmate de vogels ouder worden, terwijl de broedkansen bijna altijd hoger zijn als een vogel al een broeder was in vergelijking met die van een niet-broeder (Figuur 2d ).
Na het uitvoeren van de analyses was het tijd om de IPM te maken, waarbij de scholeksterpopulatie van de ene tijdstap naar de volgende werd gemodelleerd (Figuur 1d). De belangrijkste componenten van een IPM zijn de demografisch parameters afhankelijk van de toestand van een individu (in figuur 1d wordt dit beschreven als Z - inclusief leeftijd, broedstatus en legdatum). De IPM beschrijft hoe overleving (S) en reproductie (R) variëren tussen individuen, hoe individuen veranderen (G) van de ene tijdstap naar de volgende (bijv. veroudering, verandering in broedstatus of verandering in legdatum), en de eigenschappen van nieuwe individuen die de populatie (D) binnenkomen (ze zijn 1 jaar oud, ze hebben nog nooit gebroed en ze hebben een legdatum toegewezen gekregen die is getrokken uit een normale verdeling op basis van het gemiddelde en de SD van jonge broedvogels (<6kj)). Zoals beschreven, wilden we de levensgeschiedenis van de scholekster nauwkeurig beschrijven (Figuur 1d), dus hebben we drie broedstaten opgenomen (broeder, niet-broeder en “pre-broeder”, d.w.z. jonge exemplaren die nog niet hebben gebroed), veertig leeftijdsklassen om reproductie op latere leeftijd (>3kj) mogelijk te maken en leeftijdsspecifieke overleving. We analyseerden ook elke reproductiefase, dus in plaats van alleen het aantal geproduceerde kuikens te schatten, schatten we de kans op broeden, en voor broeders, of het nest al of niet succesvol was en hoeveel jongen er uit succesvolle nesten kwamen. Vervolgens schatten we de overleving van de jongen tot vliegvlug en daarna de overlevingskans in het eerste jaar. Hetzelfde deden we ook voor de vervolglegsels, als het eerste legsel verloren ging. De reden is dat verschillende stadia van de reproductiecyclus met verschillende bedreigingen te maken hebben, Zo zijn nesten kwetsbaarder voor overstromingen tijdens de ei-fase en de predatie van eieren of kuikens kan afhangen van het type aanwezige roofdieren (zie bijvoorbeeld Magali's paper - https://doi.org/10.1111/ddi.13457).
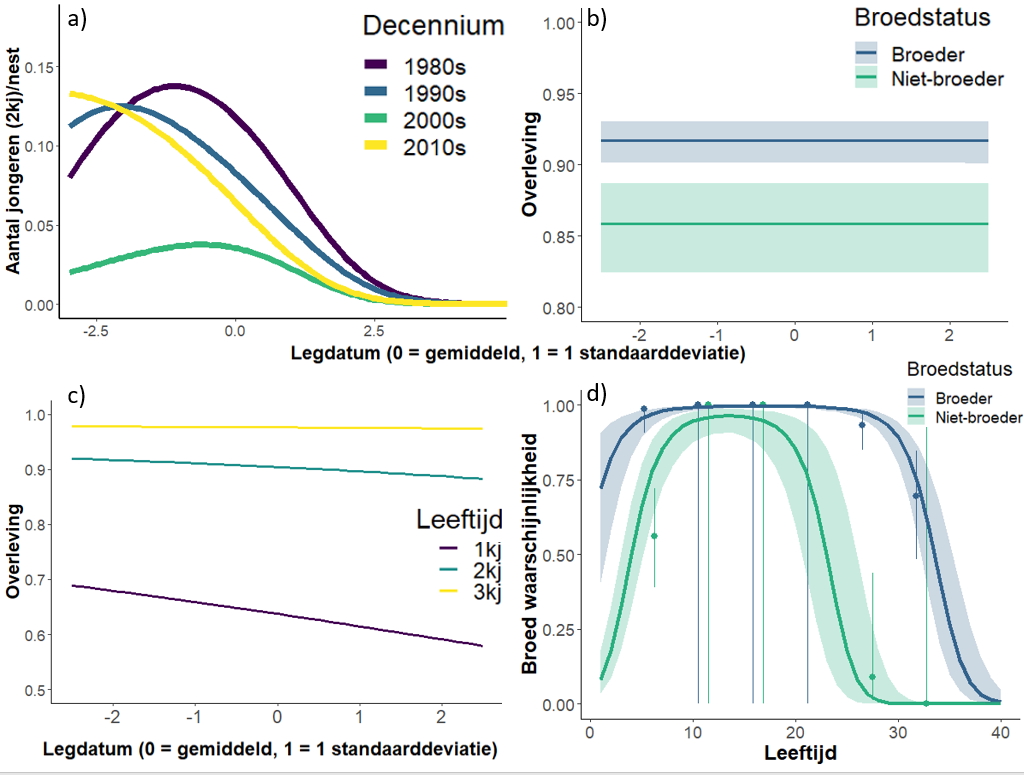
Figuur 2. Enkele voorbeelden van de verbanden die zijn vastgesteld tijdens analyses voor het populatiemodel – zie bijlage S1 van het artikel voor een veel vollediger overzicht. a) relatie tussen het gemiddeld aantal jongen dat de leeftijd van één jaar (2kj) bereikt per nest in relatie tot de legdatum van het nest (per decennium), b) Overleving van niet-broeders en broeders in relatie tot legdatum (in de laatste decennium, 2010s), c) overleving van scholeksters in de eerste drie jaar in relatie tot legdatum (in het laatste decennium, 2010s, 1kj is vliegvlug jongen), d) broedkans in relatie tot leeftijd en de huidige broedstatus (in het laatste decennium, 2010s).
Resultaten
We maakten een populatiemodel voor elk decennium, en ook een gemiddeld populatiemodel voor de hele onderzoeksperiode. In figuur 3 geven de ononderbroken lijnen de populatiegroei weer, waarbij het duidelijk is dat de scholeksterpopulatie stabiel was in de jaren tachtig van de vorige eeuw, begon af te nemen in de jaren negentig met 3% per jaar, sterk daalde in de jaren 2000-2010 met 9% per jaar alvorens te herstellen in de jaren 2010-2020 met 4% daling per jaar. Vervolgens hebben we onderzocht hoe elke demografische parameters tussen decennia varieerde door het gemiddelde van de afgelopen veertig jaar te vervangen door een parameter uit dat specifieke decennium. Om de invloed te beoordelen, moet je de gekleurde punten in figuur 3 vergelijken met de stippellijn. Punten die boven de lijn liggen, betekenen dat de demografische parameter beter was dan gemiddeld, en dus resulteerden in een hogere populatiegroei, terwijl punten onder de lijn betekenen dat de demografische parameter slechter waren dan gemiddeld en dus resulteerden in een lagere populatiegroei. Hieruit blijkt duidelijk dat de reproductiefase de grootste invloed op de groei van de populatie heeft gehad, vooral vanwege het slechte nestsucces (twee perioden: 2000-2010 en 2010-2020) en gedeeltelijk ook vanwege de overleving van de kuikens (een periode: 2000-2010). Het is echter interessant om op te merken dat de achteruitgang in de jaren negentig begon met een slechte overleving van de jonge vogels in het eerste jaar. Hoewel de reproductiesituatie in het laatste decennium (2010s) beter is dan in de jaren 2000, lijkt de overleving van volwassenen af te nemen en dit trekt de populatiegroei omlaag.
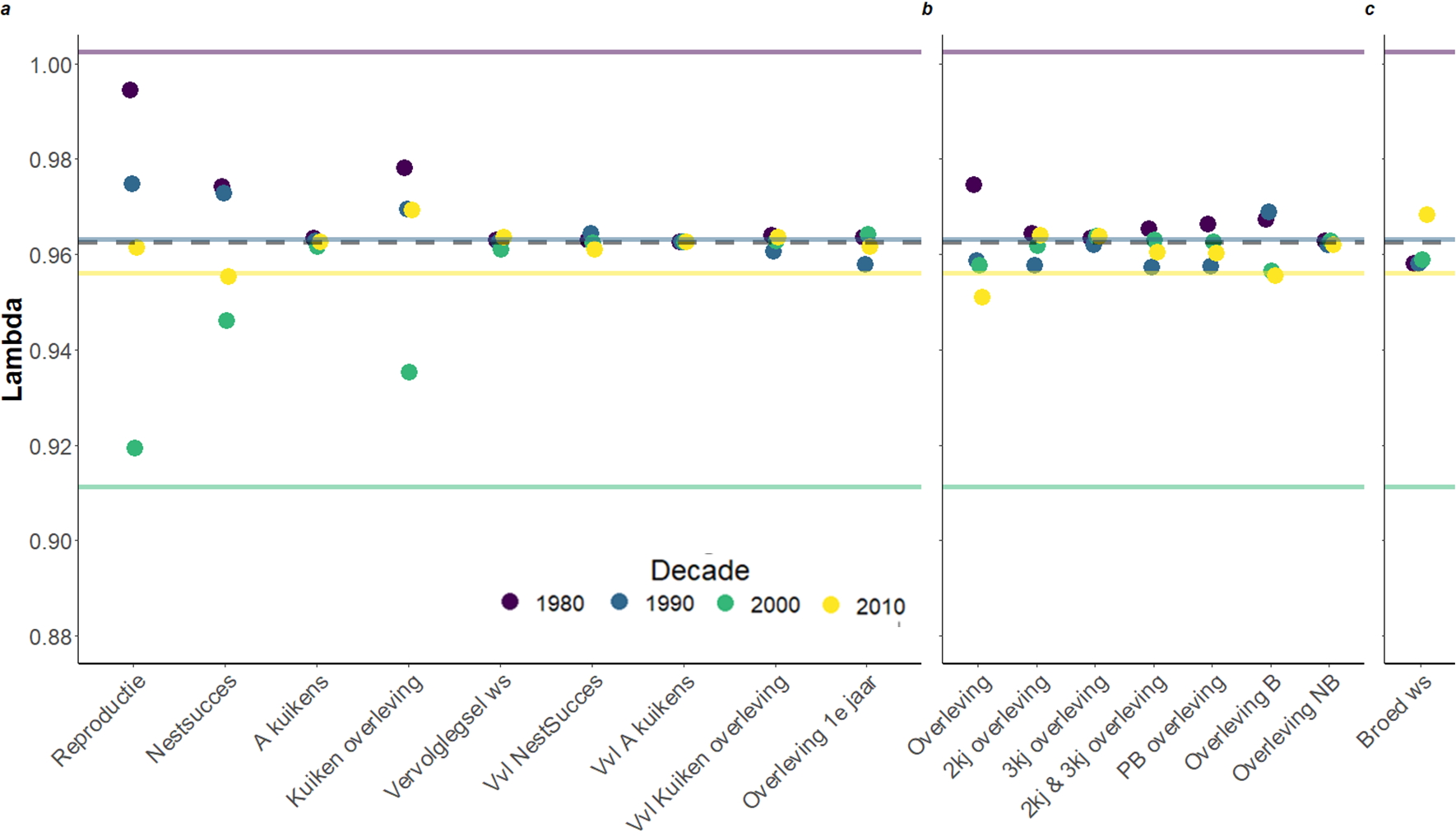
Figuur 3. Populatiegroei (lambda) van de scholekster in elk decennium (gekleurde lijnen) en de gemiddelde populatiegroei (stippellijn). De punten geven aan hoeveel de gemiddelde populatiegroei verandert als de gemiddelde demographische parameter van de laatste vier decennia wordt verwijderd en vervangen door de parameters van een bepaald decennium. De populatiegroeicijfers zijn bijvoorbeeld veel lager als de nestsuccesparameters van 2000 worden gebruikt in vergelijking met 1980. a) is alle reproductie parameters, b) is overleving parameters en c) is broed waarschijnlijkheid. A = aantal, ws = waarschijnlijkheid, Vvl = vervolglegsel, B = broeder, NB = Niet-broeder, PB = “Pre-broeder”.
Implicaties
In het artikel hebben we verschillende simulaties uitgevoerd om vast te stellen hoe de huidige populatiegroei (4% daling per jaar) verbeterd zou kunnen worden zodat de scholeksterpopulatie niet meer afneemt. We onderzochten hoeveel belangrijke demografische parameters zoals nestsucces, overleving van kuiken, of overleving van volwassen zouden moeten verbeteren. Een verbetering van een enkele demografische parameters zou geen stabiele scholeksterpopulatie opleveren, en zelfs als ze alle drie zouden terugkeren naar de niveaus die in de jaren tachtig werden waargenomen, leverde dit geen stabiele populatiegroei op, wat aangeeft waar verbeteringen nodig zijn gedurende de levenscyclus.
Een intrigerend resultaat was hoe de relatie tussen legdatum en reproductie in de loop van decennia varieerde (Figuur 2a). In de jaren tachtig hadden vogels die iets eerder dan gemiddeld tot broeden kwamen de hoogste reproductie, maar in het meest recente decennium is deze relatie veranderd, waarbij alleen de vroegste vogels het grootste succes hadden. Het probleem is dat er veel minder vogels zijn met vroege legdatums (zie inzetafbeelding van figuur 1a), wat betekent dat de reproductie van de populatie over het algemeen veel lager is, aangezien de gemiddelde vogel tegenwoordig veel minder succes heeft. In onze simulaties hebben we waargenomen dat als de gemiddelde legdatum van de populatie 10 dagen vooruit zou gaan, de scholekster populatie stabiel zou worden (als al het andere gelijk blijft). Dit roept de intrigerende vraag op wat er is veranderd op het eiland, of in de omgeving, om deze relatie te beïnvloeden - en dit is iets dat we hopen verder te onderzoeken.
Volgende stappen
Het maken van het IPM voor Schiermonnikoog was een eerste stap voor het maken van een populatiemodel voor heel Nederland. Momenteel breiden we het model uit met seizoenen (dus zomer en winter) en dus ook migratie. Het nieuwe model zal ook onderscheid maken tussen reproductie- en overlevingspercentages van kust- versus binnenvogels, vogels die overwinteren in de Waddenzee, Delta of in het buitenland, en vogels die broeden op kwelders, akkers, graslanden of in stedelijke gebieden. Op deze manier kunnen we meten hoe elke theoretische subpopulatie (d.w.z. combinatie van broedhabitat, regio en keuze van overwinteringsplaats) bijdraagt aan de totale populatie van Nederland, hoe menselijke en mlieufactoren deze demografische waarden kunnen beïnvloeden, en bijgevolg hoe we onze activiteiten kunnen optimaliseren om de achteruitgang om te buigen en een stabiele of zelfs groeiende scholeksterpopulatie te realiseren.
Publicatie:
Allen AM, Jongejans E, van de Pol M, Ens BJ, Frauendorf M, van der Sluijs M & de Kroon H. 2022. The demographic causes of population change vary across four decades in a long-lived shorebird. Ecology 103 (4), e3615.
Link












